Introduction: In the Postle lab, we are interested in the TonB system, which overcomes the limitations of outer membranes of Gram-negative bacteria. It is a virulence factor that many pathogens use to get iron from their hosts. It works by signal transduction from the energized cytoplasmic membrane to outer membrane active transporters. Three proteins in the cytoplasmic membrane, TonB, ExbB and ExbD harvest the protonmotive force of the cytoplasmic membrane and transmit it to customized transporters in the outer membrane. The model below guides our thinking about how TonB becomes energized prior to contact with transporters in Stage IV, and then recycled.
Directions: To use our interactive diagram, click on the various parts to find out more information and references. To reset the diagram, just click on the “Home” button. If you would like to access the publications related to each icon, please look below in the table for description and hyperlinks.
| Icon | Description |
|---|---|
| The proposed in vivo interactions of the TonB and ExbD periplasmic C-termini (spheres) as they are escorted through an energy transduction cycle by ExbB is shown. The ratio of TonB:ExbD:ExbB in E. coli is 1:2:7 (PMID:11967085). For a full justification of the model, see Gresock et al. 2015 PMID:26283773 |
|
 | TonB is homodimerized throughout the energy transduction cycle at or near the N-terminal transmembrane domain (PMID:26283773) |
 | ExbD forms disulfide-linked homodimers through both its N-terminal transmembrane domain (soon to be published data) and its C-terminal periplasmic domain. Some of the C-terminal homodimers are inactive suggesting that they represent a transitional state (PMID:21984795). |
| In Stage I, we have no evidence that TonB and ExbD interact (PMID:22100395), yet both require ExbB for their proteolytic stability. |
|
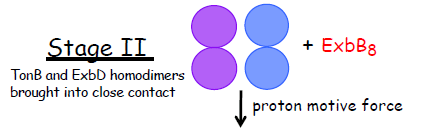 | In Stage II, ExbB tetramers coalesce into octamers through contacts with one or more unidentified proteins through their cytoplasmic domains. In so doing they bring ExbD2 and TonB2 C-termini into sufficient proximity that ExbD2 protects the N-terminal ~ 155 residues of TonB2 from proteolysis by proteinase K in spheroplasts. Stage II does not require the protonmotive force of the cytoplasmic membrane (PMID:22100395). |
 | TonB is proteolytically unstable in the absence of ExbB. ExbB crosslinks into tetramers and tetramers with ~ 85 kDa of unknown protein in vivo (PMID:21840979). |
 | ExbD is proteolytically unstable in the absence of ExbB. ExbB crosslinks into tetramers and tetramers with ~ 85 kDa of unknown protein in vivo (PMID:21840979). |
 | In Stage III, the cytoplasmic membrane protonmotive force is used to configure the relationship between the TonB2 and ExbD2 C-termini such that they exchange partners to become a pair of TonB-ExbD heterodimers. The transition between Stages II and III is reversible (PMID:22100395). |
 | Left to its own devices, the TonB C-terminus assumes structures where the aromatic residues are all buried in the interior of the protein (PMID:16039843). However genetic evidence suggests that those residues are required for binding to customized outer membrane transporters (PMID:22016747; PMID:14651622; PMID:15612934). The purpose of ExbD may be to uncover the aromatic residues to direct TonB into the correct conformation for binding to the outer membrane transporters. |
 | The TonB C-terminus binds to outer membrane transporters even when it cannot transduce energy, both in vivo (PMID:21840979) and in vitro (PMID:16741124). In Stage IV, and in the correct conformation as determined by ExbD, the TonB C-terminus energizes active transport through beta-barrel proteins with occluded lumens in a way that is still undefined. |
 | An independently acting TonB1 C-terminus transitions from a TonB-ExbD heterodimer to a TonB-FepA heterodimer (PMID:28264993) while remaining homodimerized through its N-terminus. |
 | After an energy-transduction event, signal transduction from the TonB N-terminus to C-terminus is required to restore the TonB C-terminus to its required homodimeric state such that it can again be configured by the ExbD C-terminus for another round of energy transduction. Histidine 20 in the TonB transmembrane domain is required for this re-dimerization (PMID:26283773), but unexpectedly, plays no role in utilization of the protonmotive force (PMID:21665976). |
 | Why propose a pool of depleted ExbD homodimers (ExbD2/ExbB4)? 1) There is twice as much ExbD per cell as TonB, but no evidence for a complex with four ExbDs and two TonBs. And there is a need to also account for additional ExbB proteins due to a functional ratio in cells of 2TonB:4ExbD:14 ExbB (PMID:11967085) because TonB is a homodimer throughout the energy transduction cycle (PMID:26283773). 2) ExbB has no proton-responsive residues in any of its three transmembrane domains (PMID:23603742), so its energy cannot be depleted. 3) ExbD carries the sole proton-responsive residue of the whole system in its transmembrane domain (Asp25—our soon to be published data). ExbD D25 is required for transition from Stage II to Stage III (PMID:22100395) and for correct positioning of the TonB C-terminus so that it transduces energy to an outer membrane transporter (PMID:22493017). |
 | The pool of ExbD2/ExbB4 transitions from depleted to restored and ready to participate again in Stage I. |
| This is the most speculative part of the model. ExbD is recycled, having been “used up” during an energy transduction event. One can imagine that a mystery protein acting through the cytoplasmic domain of ExbB plays a role in that event. We have looked extensively for a role for ATP hydrolysis and failed to find one. |